INTRODUCTION
In general, enzymes are proteins produced by living cells; they act as catalysts in biochemical reactions. A catalyst affects the rate of a chemical reaction. One consequence of enzyme activity is that cells can carry out complex chemical activities at relatively low temperatures.
In an enzyme-catalyzed reaction, the substance to be acted upon (the substrate = S) binds reversibly to the active site of the enzyme (E). One result of this temporary union is a reduction in the energy required to activate the reaction of the substrate molecule so that the products (P) of the reaction are formed.
Note that the enzyme is not changed in the reaction and can be recycled to break down additional substrate molecules. Each enzyme is specific for a particular reaction because its amino acid sequence is unique and causes it to have a unique three-dimensional structure. The active site is the portion of the enzyme that interacts with the substrate, so that any substance that blocks or changes the shape of the active site affects the activity of the enzyme. A description of several ways enzyme action may be affected follows.
1. Salt concentration. If the salt concentration is close to zero, the charged amino acid side chains of the enzyme molecules will attract each other. The enzyme will denature and form an inactive precipitate. If, on the other hand, the salt concentration is very high, normal interaction of charged groups will be blocked, new interactions will occur, and again the enzyme will precipitate. An intermediate salt concentration such as that of human blood (0.9%) or cytoplasm is the optimum for many enzymes.
2. pH. pH is a logarithmic scale that measures the acidity or H+ concentration in a solution. The scale runs from 0 to 14 with 0 being highest in acidity and 14 lowest. When the pH is in the range of 0-7, a solution is said to be acidic; if the pH is around 7, the solution is neutral; and if the pH is in the range of 7-14, the solution is basic. Amino acid side chains contain groups such as -COOR and -NH2 that readily gain or lose H+ ions. As the pH is lowered an enzyme will tend to gain H+ ions, and eventually enough side chains will be affected so that the enzyme's shape is disrupted. Likewise, as the pH is raised, the enzyme will lose H+ ions and eventually lose its active shape. Many enzymes perform optimumly in the neutral pH range and are denatured at either an extremely high or low pH. Some enzymes, such as pepsin, which acts in the human stomach where the pH is very low, have a low pH optimum.
3. Temperature. Generally, chemical reactions speed up as the temperature is raised. As the temperature increases, more of the reacting molecules have enough kinetic energy to undergo the reaction. Since enzymes are catalysts for chemical reactions, enzyme reactions also tend to go faster with increasing temperature. However, if the temperature of an enzyme-catalyzed reaction is raised still further, a temperature optimum is reached; above this value the kinetic energy of the enzyme and water molecules is so great that the conformation of the enzyme molecules is disrupted. The positive effect of speeding up the reaction is now more than offset by the negative effect of changing the conformation of more and more enzyme molecules. Many proteins are denatured by temperatures around 40-500C, but some are still active at 70-800C, and a few even withstand boiling.
4. Activations and Inhibitors. Many molecules other than the substrate may interact with an enzyme. If such a molecule increases the rate of the reaction it is an activator, and if it decreases the reaction rate it is an inhibitor. These molecules can regulate how fast the enzyme acts. Any substance that tends to unfold the enzyme, such as an organic solvent or detergent, will act as an inhibitor. Some inhibitors act by reducing the -S-S- bridges that stabilize the enzyme's structure. Many inhibitors act by reacting with side chains in or near the active site to change its shape or block it. Many well-known poisons such as potassium cyanide and curare are enzyme inhibitors that interfere with the active site of critical enzymes.
The enzyme used in this lab, catalase, has four polypeptide chains, each composed of more than 500 amino acids. This enzyme is ubiquitous in aerobic organisms. One function of catalase within cells is to prevent the accumulation of toxic levels of hydrogen peroxide formed as a by-product of metabolic processes. Catalase might also take part in some of the many oxidation reactions that occur in all cells.
The primary reaction catalyzed by catalase is the decomposition of H2O2 to form water and oxygen.
In the absence of catalase, this reaction occurs spontaneously, but very slowly. Catalase speeds up the reaction considerably. In this experiment, a rate for this reaction will be determined.
Much can be learned about enzymes by studying the kinetics (particularly the changes in rate) of enzyme-catalyzed reactions. For example, it is possible to measure the amount of product formed, or the amount of substrate used, from the moment the reactants are brought together until the reaction has stopped.
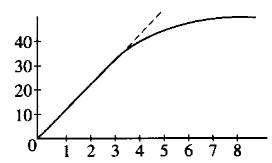
If the amount of product formed is measured at regular intervals and this quantity is plotted on a graph, a curve like the one that follows is obtained.
Figure 2.1: Enzyme Activity
|
|
Study the solid line on the graph of this reaction. At time 0 there
is no product. After 30 seconds, 5 micromoles (µmoles) have been
formed; after one minute, 10 µmoles; after two minutes, 20 µmoles.
The rate of this reaction could be given as 10 µmoles of product
formed per minute for this initial period. Note, however, that by the third
and fourth minutes, only about 5 additional ttmoles of product have been
formed. During the first three minutes, the rate is constant. From the
third minute through the eighth minute, the rate is changing; it is slowing
down. For each successive minute after the first three minutes, the amount
of product formed in that interval is less than in the preceding minute.
From the seventh minute onward, the reaction rate is very slow.
In the comparison of the kinetics of one reaction with another, a common reference point is needed. For example, suppose you wanted to compare the effectiveness of catalase obtained from potato with that of catalase obtained from liver. It is best to compare the reactions when the rates are constant. In the first few minutes of an enzymatic reaction such as this, the number of substrate molecules is usually so large compared with the number of enzyme molecules that changing the substrate concentration does not (for a short period at least) affect the number of successful collisions between substrate and enzyme. During this early period, the enzyme is acting on substrate molecules at a nearly constant rate. The slope of the graph line during this early period is called the initial rate of the reaction. The initial rate of any enzyme-catalyzed reaction is determined by the characteristics of the enzyme molecule. It is always the same for any enzyme and its substrate at a given temperature and pH. This also assumes that the substrate is present in excess.
The rate of the reaction is the slope of the linear portion of the
curve. To determine a rate, pick any two points on the straight-line
portion of the curve. Divide the difference in the amount of product formed
between these two points by the difference in time between them. The result
will be the rate of the reaction which, if properly calculated, can be
expressed as µmoles product/sec. The rate then is:
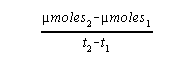 |
or from the graph, |
The rate of a chemical reaction may be studied in a number of ways,
including the following:
1. Measuring the rate of disappearance of substrate (in this example,
H2O2);
2. measuring the rate of appearance of product (in this case, O~, which
is given off as a gas); and
3. measuring the heat released (or absorbed) during the reaction.
General Procedure
In this experiment the disappearance of the substrate, H2O2,
is measured as follows:
1. A purified catalase extract is mixed with substrate (H2O2)
in a beaker. The enzyme catalyzes the conversion of H2O2
to H2O and O2 (gas).
2. Before all of the H2O2 is converted to H2O
and 02, the reaction is stopped by adding sulfuric acid (H2SO4).
The H2SO4 lowers the pH, denatures the enzyme, and
thereby stops the enzyme's catalytic activity.
3. After the reaction is stopped, the amount of substrate (H2O2)
remaining in the beaker is measured. To assay (measure) this quantity,
potassium permanganate is used. Potassium permanganate (KMnO4),
in the presence of H2O2 and H2SO4
reacts as follows:
5H2O2+2KMnO4+3H2SO4 forms K2SO4+2MnSO4+8H2O+5O2
Note that H202 is a reactant for this reaction. Once all the H2O2 has reacted, any more KMnO4 added will be in excess and will not be decomposed. The addition of excess KMnO4 causes the solution to have a permanent pink or brown color. Therefore, the amount of H2O2 remaining is determined by adding KMnO4 until the whole mixture stays a faint pink or brown, permanently. Add no more KMnO4 after this point. The amount of KMnO4 added is a proportional measure of the amount of H2O2 remaining (2 molecules of KMnO4 reacts with 5 molecules of H2O2 as shown in the equation).
EXERCISE 2A: Test of Catalase Activity
1. To observe the reaction to be studied, transfer 10 mL of 1.5% (0.44
M) H2O2 into a 50 mL beaker and add 1 mL freshly
made catalase solution. The bubbles coming from the reaction mixture are
O2, which results from the breakdown of H2O2
by catalase. Be sure to keep the freshly made catalase solution on ice
at all times.
2. The effect of boiling on enzyme activity was demonstrated in a previous
lab (Biol 121).
3. The presence of catalase in living tissue was demonstrated in a
previous lab (Biol 121).
EXERCISE 2B: The Baseline Assay
To determine the amount of H2O2 initially present
in a 1.5% solution, one needs to perform all the steps of the procedure
without adding catalase to the reaction mixture. This amount is known as
the baseline and is an index of the initial concentration of H2O2
in solution.
1. Place 10 mL of 1.5% H2O2 in a beaker.
2. Add 1 mL of H2O.
3. Add 10 mL of H2SO4 (1.0 M).
4. Mix well.
5. Remove a 5 mL sample (called an aliquot) and place it in a beaker
to assay the amount of H2O2. To do this, place the
beaker over a white surface. Use a burette to add KMnO4 dropwise
until a persistent pink or brown color is obtained.
Baseline Calculation: Baseline (mL KMnO4) = Final Burette - Initial Burette
6. Perform the baseline assay three times to obtain a meaningful average.
EXERCISE 2C: The Uncatalyzed Rate of H202 Decomposition
To determine the rate of spontaneous conversion of H2O2
to H2O and 02 in an uncatalyzed reaction, put
a small quantity of 1.5% H2O2 (about 15mL) in a beaker.
Store it uncovered at room temperature for approximately 24 hours. Repeat
steps 2-5 from Exercise 2B to determine the proportional amount of H2O2
remaining (for ease of calculation assume that 1 mL of KMnO4
used in the titration represents the presence of 1 mL H2O2
in the solution).
Calculate the amount of H2O2 spontaneously decomposed
(ml baseline - mL KMnO4).
Calculate the percent of the H2O2 that spontaneously
decomposes in 24 hours? [(mL baseline - mL 24 hours)/mL baseline] x 100
EXERCISE 2D: An Enzyme-Catalyzed Rate of H202 Decomposition
In this experiment, you will determine the rate at which a 1.5% H2O2
solution decomposes when catalyzed by the purified catalase extract.
To do this, you should determine how much H2O2 has
been consumed after 10, 30, 60, 120, 180, and 360 seconds.
If a day or more has passed since you did Exercise 2B, you must reestablish
the baseline by determining the amount of H202 present
in your 1.5% solution. Repeat the assay procedure (steps 1 through 5, page
24) and record the results below. The baseline assay should be approximately
the same value for all groups. Check with another team before proceeding.
Procedure for a Time-Course Determination
To determine the course of an enzymatic reaction, you will need to
measure how much substrate is disappearing over time. You will measure
the amount of substrate decomposed after 10, 30, 60, 120, 180, and 360
seconds. To use lab time more efficiently, set up all five of these at
the same time, and do them together. Stop each reaction at the proper time.
10 seconds
1. Put 10 mL of 1.5% H2O2 in a 50-mL glass beaker.
2. Add 1 mL of catalase extract.
3. Swirl gently for 10 seconds.
4. At 10 seconds, add 10 mL of H2SO4 (l.0M).
30, 60, 120, 180, and 360 seconds
Each time, repeat steps 1 through 4 as above, except allow the reactions
to proceed for 30, 60, 120, 180, and 360 seconds, respectively.
5. For each sample, remove a 5 mL aliquot and assay for the amount of H2O2 in the
sample. Follow the procedure as in the Baseline assay above. Should the end point be
overshot, remove another 5 mL aliquot and repeat the titration. Do not discard
any solutions until the entire lab is completed.
6. Repeat the titration three times for each sample to obtain a meaningful average.
7. Record and graph the results.
| 10 | 30 | 60 | 120 | 180 | 360 | |
| A. Baseline* | ||||||
| B. Final Reading | ||||||
| C. Initial Reading | ||||||
| D. KMnO4 consumed | ||||||
| E. H2O2 used |